* Information From OMIM
Description: Actin-binding protein, or filamin, is a 280-kD protein that crosslinks actin filaments into orthogonal networks in cortical cytoplasm and participates in the anchoring of membrane proteins for the actin cytoskeleton. Remodeling of the cytoskeleton is central to the modulation of cell shape and migration. Filamin A, encoded by the FLNA gene, is a widely expressed protein that regulates reorganization of the actin cytoskeleton by interacting with integrins, transmembrane receptor complexes, and second messengers.
» More (6) « Hide Below
Function: Vadlamudi et al. (2002) identified FLNA as a binding partner of PAK1 (OMIM:602590) in a yeast 2-hybrid screen of a mammary gland cDNA library. By mutation analysis, they localized the PAK1-binding region in FLNA to tandem repeat 23 in the C terminus, and the FLNA-binding region in PAK1 between amino acids 52 and 132 in the conserved CDC42 (OMIM:116952)/RAC (OMIM:602048)-interacting domain. Endogenous FLNA was phosphorylated by PAK1 on ser2152 following stimulation with physiologic signaling molecules. Following stimulation, FLNA colocalized with PAK1 in membrane ruffles. The ruffle-forming activity of PAK1 was found in FLNA-expressing cells, but not in cells deficient in FLNA.
Function: Androgen receptor (AR; OMIM:313700), a nuclear transcription factor, mediates male sexual differentiation. Loy et al. (2003) characterized a negative regulatory domain in the AR hinge region, which interacts with filamin A. Filamin A interferes with AR interdomain interactions and competes with the coactivator transcriptional intermediary factor-2 (TIF2; OMIM:601993) to downregulate AR function specifically. Although full-length filamin A is predominantly cytoplasmic, a C-terminal 100-kD fragment colocalized with AR to the nucleus. This naturally occurring filamin A fragment repressed AR transactivation and disrupted AR interdomain interactions and TIF2-activated AR function in a manner reminiscent of full-length filamin A, raising the possibility that the inhibitory effects of cytoplasmic filamin A may be transduced through this fragment, which can localize to the nucleus and form part of the preinitiation complex. This unanticipated role of filamin A added to the evidence for the involvement of cytoskeletal proteins in transcription regulation.
Function: Mutation in the X-linked FLNA gene can cause the neurologic disorder periventricular heterotopia (OMIM:300049). Although periventricular heterotopia is characterized by a failure in neuronal migration into the cerebral cortex with consequent formation of nodules in the ventricular and subventricular zones, many neurons appear to migrate normally, even in males, suggesting compensatory mechanisms. Sheen et al. (2002) showed that, in mice, Flna mRNA was widely expressed in all brain cortical layers, whereas a homolog, Flnb (OMIM:603381), was most highly expressed in the ventricular and subventricular zones during development. In adulthood, widespread but reduced expression of Flna and Flnb persisted throughout the cerebral cortex. Flna and Flnb proteins were highly expressed in both the leading processes and somata of migratory neurons during corticogenesis. Postnatally, Flna immunoreactivity was largely localized to the cell body, whereas Flnb was localized to the soma and neuropil during neuronal differentiation. The putative Flnb homodimerization domain strongly interacted with itself or the corresponding homologous region of Flna, as shown by yeast 2-hybrid interaction. The 2 proteins colocalized within neuronal precursors by immunocytochemistry, and the existence of Flna-Flnb heterodimers could be detected by coimmunoprecipitation. Sheen et al. (2002) suggested that FLNA and FLNB may form both homodimers and heterodimers, and that their interaction could potentially compensate for the loss of FLNA function during cortical development within patients with periventricular heterotopia.
Function: Using a yeast 2-hybrid screen, Grimbert et al. (2004) identified FLNA as a binding partner for both CMIP (OMIM:610112) and its truncated isoform, TCMIP. Coimmunoprecipitation analysis confirmed the interactions. Immunofluorescence microscopy demonstrated homogeneous colocalization of CMIP and FLNA in the cytoplasm, but restriction of TCMIP/FLNA colocalization to points of intercellular contact. Western blot analysis showed increased FLNA expression in patients with relapse of minimal change nephrotic syndrome, a glomerular disease thought to result from abnormal T-cell activation. Grimbert et al. (2004) proposed that FLNA and CMIP/TCMIP interact in a T-cell signaling pathway.
Function: Using proteomic approaches, Thelin et al. (2007) showed that FLNA associates with the extreme CFTR (OMIM:602421) N terminus. Cell studies revealed that filamin tethers plasma membrane CFTR to the underlying actin network, stabilizing CFTR at the cell surface and regulating the plasma membrane dynamics and confinement of the channel. In the absence of filamin binding, CFTR is rapidly internalized from the cell surface, where it accumulates prematurely in lysosomes and is ultimately degraded.
Function: Using yeast 2-hybrid analysis and protein pull-down assays, Jimenez-Baranda et al. (2007) showed that the human immunodeficiency virus (HIV)-1 (see OMIM:609423) receptor CD4 (OMIM:186940) and the HIV-1 coreceptors CCR5 (OMIM:601373) and CXCR4 (OMIM:162643) interacted with FLNA, which regulated clustering of the HIV-1 receptors on the cell surface. Binding of HIV-1 gp120 to the receptors induced transient cofilin (see CFL1; OMIM:601442) phosphorylation inactivation through a RHOA (OMIM:165390)-ROCK (see OMIM:601702)-dependent mechanism. Blockade of FLNA interaction with CD4 and/or the coreceptors inhibited gp120-induced RHOA activation and cofilin inactivation. Jimenez-Baranda et al. (2007) concluded that FLNA is an adaptor protein that links HIV-1 receptors to the actin skeleton remodeling machinery, possibly facilitating virus infection.
* References
[PubMed ID: 12477932] Strausberg RL, Feingold EA, Grouse LH, Derge JG, Klausner RD, Collins FS, Wagner L, Shenmen CM, Schuler GD, Altschul SF, Zeeberg B, Buetow KH, Schaefer CF, Bhat NK, Hopkins RF, Jordan H, Moore T, Max SI, Wang J, Hsieh F, Diatchenko L, Marusina K, Farmer AA, Rubin GM, Hong L, Stapleton M, Soares MB, Bonaldo MF, Casavant TL, Scheetz TE, Brownstein MJ, Usdin TB, Toshiyuki S, Carninci P, Prange C, Raha SS, Loquellano NA, Peters GJ, Abramson RD, Mullahy SJ, Bosak SA, McEwan PJ, McKernan KJ, Malek JA, Gunaratne PH, Richards S, Worley KC, Hale S, Garcia AM, Gay LJ, Hulyk SW, Villalon DK, Muzny DM, Sodergren EJ, Lu X, Gibbs RA, Fahey J, Helton E, Ketteman M, Madan A, Rodrigues S, Sanchez A, Whiting M, Madan A, Young AC, Shevchenko Y, Bouffard GG, Blakesley RW, Touchman JW, Green ED, Dickson MC, Rodriguez AC, Grimwood J, Schmutz J, Myers RM, Butterfield YS, Krzywinski MI, Skalska U, Smailus DE, Schnerch A, Schein JE, Jones SJ, Marra MA, Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proc Natl Acad Sci U S A. 2002 Dec 24;99(26):16899-903. Epub 2002 Dec 11.

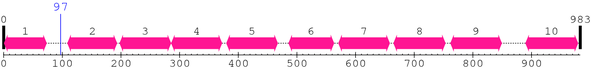

 Thr
Thr
 Sequence conservation (by blast)
Sequence conservation (by blast)